
Ch 12: extract 7
Is anything completely purposeless?

I’m very much hoping that no potential readers think they need to have a degree in biology to see why this chapter we are exploring, Chapter 12, seems to me of some considerable philosophical importance. Hardly. It requires almost no prior detailed knowledge, since nothing is taken for granted. But what it reveals about the nature of living organisms is, I believe, of the first importance, whether you are interested in biology or not. And hence my - seemingly rather oddly - singling out this chapter from a 30-chapter book, that tries to get as close as we can, from many different sources, to the nature of reality itself. What is to me beautiful and confirming is that, wherever we look - whether to the revelatory capacity of great art, to physics, philosophy, or biology, to the science of mind and brain, and to the great narrative traditions and myths embodying hard-earned wisdom in the spiritual traditions of people all over the world since time immemorial - we see a coherent picture emerge. It is that picture that I have wished to put to the proof at length in my books, and ultimately convey to my readers. A picture of a world more wonderful than we have allowed ourselves to admit seeing, or even to see at all. If there is meaning to life, then nothing can be more important than the quest to encounter it more clearly. And if there isn’t, and you are sure of that, relax. It’ll all be over soon enough.
The ‘dreadful’ question of purpose
The problem biology faces, but physics doesn’t, is that the phenomena it is trying to explain require it to ask not one question, but two. This has been succinctly pointed out by George Gaylord Simpson:
‘How?’ is the typical, and only meaningful, question in the physical sciences. But biology can and must go on from there. Here, ‘What for?’ – the dreadful teleological question – not only is legitimate but also must eventually be asked of every vital phenomenon … In biology, then, a second kind of explanation must be added to the first, or reductionist, explanation made in terms of physical, chemical, and mechanical principles. This second form of explanation, which can be called compositionist in contrast with reductionist, is in terms of the adaptive usefulness of structures and processes to the whole organism.[1]
Biology has a problem with teleology. In J.B.S. Haldane’s oft-quoted words, ‘teleology is like a mistress to a biologist; he cannot live without her but he’s unwilling to be seen with her in public’.[2] However, avoiding teleology results in vacuity, as the evolutionary biologist Ernst Mayr trenchantly observed:
Teleonomic statements have often been maligned as stultifying and obscurantist. This is simply not true. Actually, the nonteleological translation is invariably a meaningless platitude, while it is the teleonomic statement which leads to biologically interesting inquiries …’ [3]
Mayr is thinking principally of those scientists – and they exist – who want us to avoid making specific teleological observations of behaviour, such as ‘a turtle comes ashore in order to lay her eggs’, by saying ‘a turtle comes ashore and lays her eggs’. This seems like a hangover from the bad old days of behaviourism, where all inner life is denied, even in humans. Yet, as Oxford professor of physics Andrew Steane says, ‘the structure of science is not one which dismisses the higher-level categories and “explains them away”. That is not what “scientific” means.’ Even Jacques Monod, an ardent reductionist, who is said (probably wrongly) to have said that ‘nature does not have any intention or goal’, wrote:
one of the fundamental characteristics common to all living beings without exception [is] that of being objects endowed with a purpose or project, which at the same time they show in their structure and execute through their performances … Rather than reject this idea (as certain biologists have tried to do), it must be recognised as essential to the very definition of living beings.[4]
Are we talking of the purpose of a single cell, or a single organism, or an evolutionary strain to which the organism belongs? All three: separating them out is to a large extent artificial, since they are different aspects of one process. At the macro level, according to palaeobiologist Simon Conway Morris, the development of the eye – improbably complex as it is – has happened independently 10 times (the camera eye six times and the compound eye four times) over the course of evolution.[5] At an intermediate level, a strain of flies, genetically manipulated so as to prevent eye formation, within a few generations produces eyes.[6] At the micro level, a single cell of the creature is capable of reacting to unforeseeable circumstances so as to preserve its function in the whole organism, in ways that suggest intelligence.
In a parallel situation, where the same mechanisms of echolocation in bats and whales were found to have evolved entirely separately, and depending in either case on the same complex co-occurring genetic changes, the first author, Dr Joe Parker of Queen Mary’s School of Biological and Chemical Sciences, London, commented: ‘We had expected to find identical changes in maybe a dozen or so genes but to see nearly 200 is incredible. We know natural selection is a potent driver of gene sequence evolution, but identifying so many examples where it produces nearly identical results in the genetic sequences of totally unrelated animals is astonishing.’[7]
Importantly, the idea of teleology, or purpose, does not entail determination: no prior plan, involving a sequence of predetermined steps to bring it about, is required. It is exactly that that is avoided. For example, a young woman may purpose to be a mother, and this purpose may be the most important thing in her life, yet it does not entail a series of steps, nor can its path be predicted. Nonetheless, if it is important, it will tend to happen. A purpose here is not a plan. It is a tendency inseparable from – woven into, as it were, the fabric of – a life, which leaves all the detail, and even the final outcome, undetermined. Like ‘at-tending’, tending alters the world and makes it what it is for us. The root, in either case, is Latin tendere, to reach out to something – something, then, in front or above. Nothing to do with being pushed, mechanically, from behind or below.
The example can be usefully taken further. It would be easy to see how to prevent the purpose succeeding: for it to have a chance of succeeding, the woman must put herself in the way of its coming about. However, that leaves when, where, and with whom, completely undetermined and undeterminable. By doing or not doing certain things, the desired outcome can be made more likely. Both at the individual and universal level, purpose exists through what one might call a shaping of the probability landscape. Thus we tend towards certain ends, while leaving exactly what happens, and how, in any one case, undetermined.
To move from a focus on things as the ultimately important constituents of reality, to one in which processes are the important entities, requires a shift in our perception of space. We relinquish the idea of causation as strictly ‘bottom up’, from parts to wholes: we come to accept that whole systems contribute to the properties of their parts.[8] The first is the only one that makes sense to the left hemisphere; in its view the whole can be understood only if we understand the parts. It takes the right hemisphere to understand that the parts can be understood only in the light of the whole.
But it also involves a shift in our perception of time. It is not just that chains of causation work from the past towards the future: the future, in the sense of internalised potential, pattern or telos – may be as important a driver in the emergence of phenomena as the past. As Joseph Woodger says, ‘in regard to the explanation, a knowledge of the outcome of a biological process is often more illuminating than knowledge of what went before.’[9]
Form in living creatures is, despite appearances, another aspect of the question about purpose: both imply a tendency, a direction of intent, an overall attractive outline. Some ‘sense’ of this, as Woese and McClintock would say, is what enables the organism to repair, or correct, itself. In other words, to correct itself is an inclination towards a form that is its goal, towards a goal that is its form.
The purpose I am talking about is nothing extrinsic, but rather intrinsic potential that is fulfilled within a process as the process unfolds. ‘Latent within the polymeric sequence of amino acids that constitutes the enzyme’s primary structure’, writes Stein, ‘is directionality and potential for its correct folding into a catalyst of remarkable power and selectivity.’[10]The forms that are required in the enzyme, in the cell, in the tree, may not be achieved by pushing from behind, but by drawing from in front, towards certain ends, certain conformations, certain attractors, much as a valley ‘draws’ water that falls evenly on the surrounding hills to one clear end: the mouth of just one river. There is stored attractive energy in the formation of the landscape.[11]
So a system, including a cell or a multicellular organism, can be free and undetermined, yet despite the occurrence of random events, exhibit patterned and purposeful behaviour. To quote Paul Weiss:
Small molecules go in and out, macromolecules break down and are replaced, particles lose and gain macromolecular constituents, divide and merge, and all parts move at one time or another, unpredictably, so that it is safe to state that at no time in the history of a given cell, much less in comparable stages of different cells, will precisely the same constellation of parts ever recur . . . [yet] their joint behavior converges upon a nonrandom resultant, keeping the state of the population as a whole relatively invariant.[12]
Mutations have been supposed to be random with respect to fitness: those that bring benefit persist, those that do not die out. But such a claim requires clarity about how to judge fitness. This is far from straightforward. There is no clear way of determining fitness except in that a thing does survive, which is circular. The geneticist Theodosius Dobzhansky – the same man who said that ‘nothing in biology makes sense except in the light of evolution’ – wrote that no biologist ‘can judge reliably which “characters” are useful, neutral, or harmful in a given species’ (see note).[13]
Geneticist Henrik Kaessmann points to the recent discovery of protein-coding genes being composed de novo (‘from scratch’), ie, from non-protein-coding genomic sequences altogether unrelated to pre-existing genes or transposable sequences. Such creation was widely thought to be virtually impossible: indeed, according to the famous French biologist François Jacob the probability for creation of new protein-coding genes de novo by random processes ‘is practically zero’.[14] And yet ‘recent work has uncovered a number of new protein-coding genes that apparently arose from previously noncoding (and nonrepetitive) DNA sequences’.[15] What is more, according to another research team, there is ‘accumulating evidence that de novo evolution of genes from noncoding sequences could have an important role’ in a class of genes representing ‘up to one third of the genes in all genomes’.[16]
As Talbott points out, there has, until recently, been what looks like a doctrinaire assumption of randomness projected onto ‘a descriptive biological literature that, from the molecular level on up, remains almost nothing but a documentation of the meaningfully organized, goal-directed stories of living creatures.’ And he continues:
Here, then, is what the advocates of evolutionary mindlessness and meaninglessness would have us overlook. We must overlook, first of all, the fact that organisms are masterful participants in, and revisers of, their own genomes, taking a leading position in the most intricate, subtle, and intentional genomic ‘dance’ one could possibly imagine. And then we must overlook the way the organism responds intelligently, and in accord with its own purposes, to whatever it encounters in its environment, including the environment of its own body, and including what we may prefer to view as ‘accidents’. Then, too, we are asked to ignore not only the living, reproducing creatures whose intensely directed lives provide the only basis we have ever known for the dynamic processes of evolution, but also all the meaning of the larger environment in which these creatures participate – an environment compounded of all the infinitely complex ecological interactions that play out in significant balances, imbalances, competition, cooperation, symbioses, and all the rest, yielding the marvellously varied and interwoven living communities we find in savannah and rainforest, desert and meadow, stream and ocean, mountain and valley. And then, finally, we must be sure to pay no heed to the fact that the fitness, against which we have assumed our notion of randomness could be defined, is one of the most obscure, ill-formed concepts in all of science.[17]
In an earlier chapter I suggested that attempting to break down the Gestalt of the cat into a host of useful separate adaptations that enable it to hunt well, and then being faced with the question of what started the process and what led to what, creates its own problems. It is the linear, serial processing which the left hemisphere imposes on what is a complex whole. Similarly,
the evolution of complex adaptations requires coordination between an organism’s various developmental systems. For example, the adaptive evolution of tetrapod limb structures requires coordination between the development of bone, muscle, nervous, circulatory, and integumentary systems (at least). If each system had to wait for a fortuitous mutation in order to produce the appropriate accommodation, complex evolutionary adaptations might never arise.[18]
There are aspects of the evolutionary process, as Scott Turner puts it, that ‘make little sense without intentionality of some form guiding them’.[19]
To return to purposive behaviour in an individual organism. According to Ford, ‘our conviction that these phenomena become manifest only through cell communities is a fundamental misconception. Ingenious, perceptive and intelligent behaviour is apparent in a single living cell.’[20] No-one who has ever done such a homely thing as look at drops of pond water under a microscope would ever fall for the idea that purposive, intelligent behaviour is not exhibited at the level of a single cell. ‘Single cells truly can consider their options’, writes Ford, ‘and modify their responses in the light of contingency’:
Under the microscope, we can observe a predatory single-celled ciliated microorganism as it inspect its prey from a distance. The ciliate can then select a specific microbial cell, pause, and then swoop upon its prey and capture it within a second. The coordination of this activity is similar to watching a cat catch a sparrow on the lawn, yet it is done within the confines of a single cell. Living cells are smart: the ciliate Spirostomum has been shown to possess memory.[21] It can be trained.[22]
Describing how cells often act apparently as individuals, so as to ‘seek out and destroy an invading pathogen without external mediation’, if necessary altering conventional behaviour to pursue it, and ‘maintain pursuit relentlessly until the organism is consumed and eliminated’, Ford reminds us that the human polymorph, a white blood cell that forms an important part of the body’s immune defences,
is an amoeba, although a constituent part of our bodies. This is a cell with a mind of its own. Conceptualising the human body as a cooperative community of essentially autonomous entities gives us a more reasonable understanding than our modern models, which see the body as a collection of mechanical organs enclosed in skin.[23]
Such behaviour, ‘including the continual and carefully choreographed machinations of mitochondria, the endless migration of granules and voiding of vacuoles, the conduction of discrete particles in two-way streams of cytoplasm like traffic on a highway, the meticulous changes of position of the nucleus in diatoms during division, and the cautious inspection of prey by a predatory ciliate’,[24] leads us to see something inexpressibly more complex than we can explain, never mind understand. ‘Our failure to appreciate the majesty of the cell is astonishing, and our sublime confidence that we are close to an understanding [of] it is an embarrassment’.[25]
Slime moulds are amoeba-like organisms found on moist decaying organic matter. Clearly they have no neurones, each consisting of just a single, giant cell. They are, however, capable of behaviour that looks intelligent. They are attracted to one another and fuse, creating a plasmodium, which may be up to several metres long – the slime mould that is visible to human eyes. This plasmodium is essentially a flat, liquid-filled sac, but it behaves like an organised colony:
Every part rhythmically expands and contracts, pushing around the fluid inside. If one part of the plasmodium touches something attractive, like food, it pulses more quickly and widens. If another part meets something repulsive, like light, it pulses more slowly and shrinks. By adding up all of these effects, the plasmodium flows in the best possible direction without a single conscious thought. It is the ultimate in crowdsourcing.[26]
These plasmodia can make decisions, comparing different options (for example, food sources) and selecting the best one: the plasmodium touches all the potential meals and then either ‘decides’ to move towards one, or splits itself among many. It can even balance its diet.[27] A mould can forage while making decisions with more than one objective, and it can be persuaded experimentally, as can humans, to behave irrationally at times.[28] It even turns out that moulds make better decisions if they are not rushed.[29]
Plasmodia regularly break out of Petri dishes in laboratories, and can escape from traps and solve mazes.[30] Slime mould colonies can learn to avoid certain paths that lead to a noxious stimulus, and moulds that have so learnt can transfer this memory of the adaptive response by cell fusion to new colonies that have never encountered the noxious stimulus. Even in cases where three out of four moulds in the fused organism were ‘naïve’ and had had no opportunity to learn, the information was transferred to the resulting fused slime mould from the one that had had the noxious encounter.[31]
Does a slime mould have awareness? If it does, and I suspect it does, the awareness cannot be much like ours. It does have memories and would seem to act intentionally upon them. According to Scott Turner, ‘arguably, the roots of our own intentionality permeate deeply through the living world: the living world may be rife with intentionality.’[32] I believe this to be a reasonable deduction, and will have more to say about it in Chapter 27.
It seems to me that an explanation in terms of a whole that follows attractive formal patterns or fields – Gestalten – is better than an explanation in terms of chains of causation of parts. That we cannot as yet ‘see’ or measure such patterns or fields should not be a definitive argument against entertaining the possibility of their presence. Many physical forces cannot be directly visualised or measured: we posit their existence only on the basis of their apparent effects.
Such is the case with a magnetic field. We visualise it through its effects on metallic bodies. Below is a simple illustration from D’Arcy Thompson, showing cytokinesis (a phase of cell reproduction) in a fish egg. The similarity to iron filings in the magnetic field of a bar magnet is so striking that it has been remarked on for 150 years.[33]
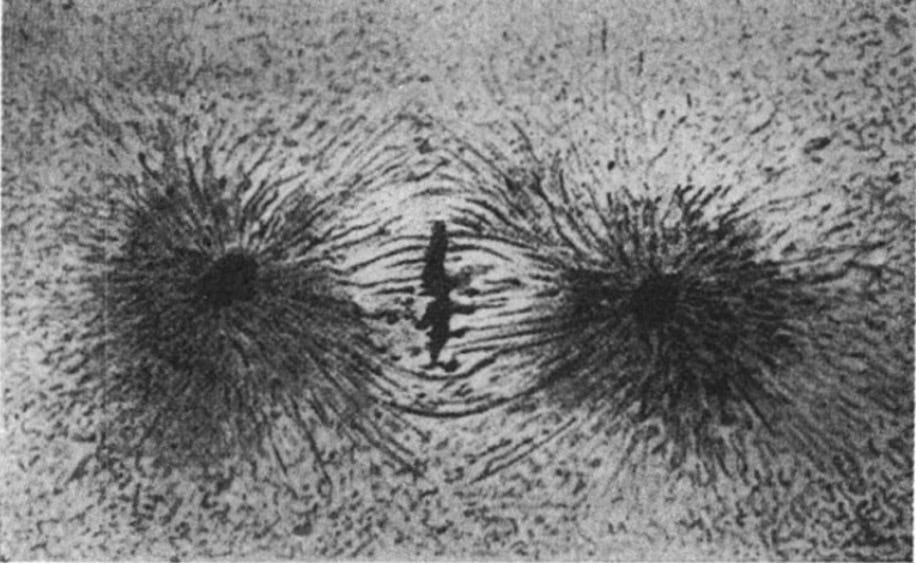
‘Caryokinetic figure in a dividing cell (or blastomere) of a trout’s egg. After Prenant, from a preparation by Prof Bouin.’ (from Thompson 1992, 299)
Tomorrow, what we can learn from desperate attempts to save the machine model …
[1] Simpson 1963 (emphasis added).
[2] Quoted in Mayr 1976 (392). A similar formulation is also attributed to the German physiologist Ernst Wilhelm von Brücke (see Cannon 1945, 108).
[3] Mayr 1989 (55).
[4] Monod 1977 (20). I cannot find a source for ‘nature does not have any intention or goal’. It is the quote from Monod most widely repeated across the internet: the sources are always unknown or non-existent. It may be a case of an unfounded ‘truth’ becoming ‘real’ through simple repetition. Claims are made (eg, on the Wikipedia page for Monod, though it gives no page reference and I suspect may not have been fully researched) that it might come from Chance and Necessity, a book I first read in my teens. However, I cannot find it there. Tellingly, it is not included, despite its currency, on Monod’s Wikiquote page. I welcome any hints from readers.
[5] Conway Morris 2005. See also Parker, Tsagkogeorga, Cotton et al 2013.
[6] See p* above.
[7] Michelson 2013.
[8] Dupré 2017a.
[9] Woodger 1929 (432).
[10] Stein 2004.
[11] This is Conrad Hal Waddington’s idea of ‘chreodes’, which I will return to in Chapter 27.
[12] Weiss 1962.
[13] Dobzhansky 1975. For an overview of this question, see Talbott 2011b: ‘Organism-environment relations present us with so much complexity, so many possible parameters to track, that, apart from obviously disabling cases, there is no way to pronounce on the significance of a mutation for an organism, let alone for a population or for the future of the species … If reduced fitness can be on the path toward higher fitness, and if the environment for which the organism is supposed to be fit is itself a modifier of the organism’s fitness, then to what solid and stable ground do we anchor our idea of fitness?’ Indeed biologists and philosophers, as Talbott points out, have yet to provide an adequate interpretation of fitness (Millstein & Skipper 2007); ‘no concept in evolutionary biology has been more confusing’ than that of fitness (Ariew & Lewontin 2004). ‘Yet the neo-Darwinian theory of natural selection hinges, in its “status … as empirical science,” upon a reasonable understanding of what fitness means (Bouchard & Rosenberg 2004) … Not only do we have great difficulty locating meaningless chance in the context of the actual life of organisms; it now turns out that the one outcome with respect to which randomness of mutation is supposed to obtain – namely, the organism’s fitness – cannot be given any definite or agreed-upon meaning, let alone one that is testable’ (Talbott op cit, 46–52).
[14] Jacob 1977. Here again I am indebted to Stephen Talbott.
[15] Kaessmann 2010.
[16] Tautz & Domazet-Lošo 2011 (emphasis added).
[17] Talbott op cit (52: emphasis in original).
[18] Pfennig, Wund, Snell-Rood et al 2007.
[19] Turner 2007 (224).
[20] Ford 2017.
[21] Beck 1975.
[22] Ford op cit.
[23] ibid.
[24] ibid.
[25] ibid.
[26] Yong 2010.
[27] Bonner 2010.
[28] Nicolis, Zabzina, Latty et al 2011; Latty & Beekman 2011a.
[29] Latty & Beekman 2011b; Beekman & Latty 2015.
[30] Reid, Latty, Dussutour et al 2012.
[31] Vogel & Dussutour 2016. See also Yong 2016.
[32] Turner 2007 (226).
[33] According to Thompson, it was first commented on (in the egg of a medusa) by Hermann Fol (1873, 476), the Swiss zoologist and father of modern cytology; then by at least six others that he names. I mention Driesch at p * above.
Henri Bergson wrote: “That life is a kind of mechanism I cordially agree. But is it the mechanism of parts artificially isolated within the whole of the universe, or is it the mechanism of the real whole? The real whole might well be, we conceive, an indivisible continuity. The systems we cut out within it would, properly speaking, not then be parts at all; they would be partial views of the whole. And, with these partial views put end to end, you will not make even a beginning of the reconstruction of the whole, any more than, by multiplying photographs of an object in a thousand different aspects, you will reproduce the object itself. So of life and of the physico-chemical phenomena to which you endeavor to reduce it.” (Bergson 1907/1944, 36)
Maurice Merleau-Ponty wrote: “[If] man really becomes the manipulandum he takes himself to be, we enter into a cultural regimen where there is neither truth nor falsity concerning man and history, into a sleep or a nightmare, from which there is no awakening. (Merleau-Ponty 1961/2004, 292)
Mazvita Chirimuuta provides these, and many other striking quotes from the rich vein of process philosophy, in her exemplary paper “The Reflex Machine and the Cybernetic Brain: The Critique of Abstraction and its Application to Computationalism”
https://www.pure.ed.ac.uk/ws/portalfiles/portal/163478123/Chirimuuta_forthcoming_Reflex_machine_penultimate.pdf
I was struck while reading this by the deep resonance between your framing of teleology in biology and Alfred Adler’s use of teleology in psychology. Though working in different domains, both perspectives seem to recognize a similar motivational essence: that life unfolds not as a reaction to what has been, but in response to what calls us forward.
Adler spoke of the individual psyche as guided by unconscious goals or imagined ideals. Not as rigid plans, but as directional tendencies shaping behavior from within. Much like your description of organisms tending toward form or coherence, this vision of purpose is open-ended, relational, and emergent.
It’s powerful to see this convergence, a shared recognition that purpose is not imposed from outside, nor reducible to mechanism. Rather, it is something woven into the living process itself, shaping, drawing, and inviting all at once.
Thank you for this beautiful and thought-provoking exploration.